In Part I and Part II of this series, I discussed genetic algorithms and why we might want to create artificial machines that begin life as a single cell, and develop into networks of identical communicating cells. In this post, I want to begin describing a model that works along these lines.
The model is a highly stylized and simplified cartoon of biological multicellular organisms. It my attempt to make the simplest model possible that captures the essence of what is happening in biology. So understand that biology is more complicated than this model; but the goal is a model stripped down to those essential elements that cannot be taken away if one wants something that looks like life. Thus, the model is proposed in the spirit of the Ising model of magnets in statistical physics; the simplest model that captures the general behavior we are looking for.
The first question is what do we want our machine (or “circuit” or “network” or “organism”; I will use these terms interchangeably) to do? As is quite conventional in hardware design, I will presume the organism receives some input signals from the world, and it is supposed to produce some desired output signal, which depends on the inputs it has received at the current and previous times. Thus, the circuit should in general be capable of creating memories, that lets it store something about previous inputs.
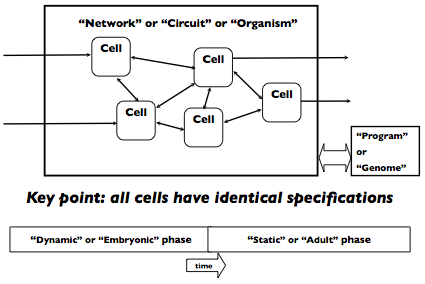
The organism begins its life as a single cell, and then has two phases in its life, a dynamic “embryonic” phase and a static “adult” phase. During the embryonic phase, the cells in the organism can undergo developmental events, primarily cell duplication, but also perhaps cell death or cell relocation, to sculpt out the final network of communicating cells. After the embryonic phase is complete (say after a fixed amount of time has passed, or some signal is generated by the circuit) the adult phase is entered. The network is static in structure during the adult phase. It is during the adult phase that the network can be tested to see whether it properly computes the desired input-output function. The figure above is a pictorial representation of the model that hopefully makes clear what I have in mind.
Each of the cells in the network will have an internal structure, defined primarily by “logic units” which send signals to each other. The computations performed by the organism will simply be the computations performed by the logic units inside of its cells. The details of what the logic units do, and how they are connected to each other, is specified by a “genome” or “program” for the organism.
Look at the figure below for a peek inside an individual cell in the model. Each cell will have an identical set of logic units, with identical connections between the logic units.
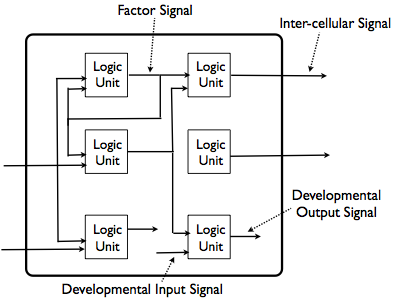
The logic units compute an output according to some fixed function of their inputs. They transmit that output after some delay, which is also part of their fixed function. The output of one logic unit will be the input of another; they send “signals” to each other.
These signals are of various types (see the above figure). The first type of signal, called a “factor signal,” will always go from a logic unit to another logic unit in the same cell. The second type of signal, called an “inter-cellular signal,” will always go from a logic unit to a logic unit in a different cell. The third type of signal, called a “developmental output signal,” will not actually go to another logic unit, but will be a signal to the cell development apparatus to perform some important development event, such as duplication or programmed cell death. Finally, the fourth type of signal, called a “developmental input signal,” will be used by the cell development apparatus to signal that some type of cell development event has occurred, and will serve as an input to logic units.
Remember that initial cell (the “fertilized egg”) will need to have a set of logic units that enable it to automatically create the adult network, so it must effectively contain the instructions for development as well as for the adult circuit. It might seem hard to imagine that this can work, but it can. In the next post in this series, I will discuss in more detail the process of development in this model, and then we will be in position to look at some interesting multicellular circuits that I have designed.
If you don’t want to wait, you can visit this page to find a PDF and a PowerPoint version of a talk I gave on the subject at a conference in Santa Fe in May 2007, although unfortunately, it might be hard to decipher without my explanation…
















{kind=link}